19.4 Independent Assortment
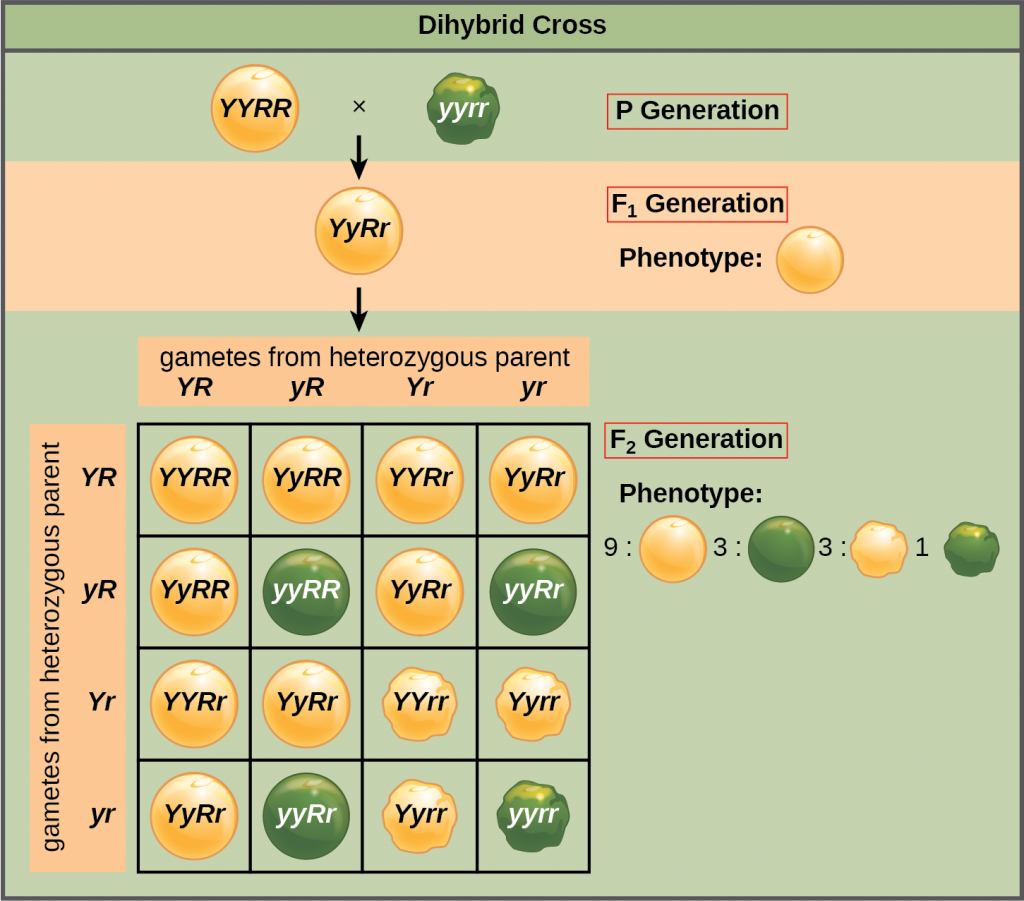
Mendel’s law of independent assortment states that genes do not influence each other with regard to the sorting of alleles into gametes, and every possible combination of alleles for every gene is equally likely to occur. The independent assortment of genes can be illustrated by the dihybrid cross, a cross between two true-breeding parents that express different traits for two characteristics.
Consider the characteristics of seed color and seed texture for two pea plants, one that has green, wrinkled seeds (yyrr) and another that has yellow, round seeds (YYRR). Because each parent is homozygous, the law of segregation indicates that the gametes for the green/wrinkled plant all are yr, and the gametes for the yellow/round plant are all YR. Therefore, the F1 generation of offspring all are YyRr .
For the F2 generation, each gamete receives either an R allele or an r allele and either a Y allele or a y allele. The law of independent assortment states that a gamete into which an r allele sorted would be equally likely to contain either a Y allele or a y allele. Thus, there are four equally likely gametes that can be formed when the YyRr heterozygote is self-crossed, as follows: YR, Yr, yR, and yr.
Arranging these gametes along the top and left of a 4 × 4 Punnett square gives us 16 equally likely genotypic combinations. From these genotypes, we see a phenotypic ratio of 9 round/yellow:3 round/green:3 wrinkled/yellow:1 wrinkled/green. These are the offspring ratios we would expect, assuming we performed the crosses with a large enough sample size.
Because of independent assortment and dominance, the 9:3:3:1 dihybrid phenotypic ratio can be collapsed into two 3:1 ratios, characteristic of any monohybrid cross that follows a dominant and recessive pattern. Ignoring seed color and considering only seed texture in the above dihybrid cross, we would expect that three quarters of the F2 generation offspring would be round, and one quarter would be wrinkled. Similarly, isolating only seed color, we would assume that three quarters of the F2 offspring would be yellow and one quarter would be green. The sorting of alleles for texture and color are independent events, so we can apply the product rule.
Therefore, the proportion of round and yellow F2 offspring is expected to be (3/4) × (3/4) = 9/16, and the proportion of wrinkled and green offspring is expected to be (1/4) × (1/4) = 1/16. These proportions are identical to those obtained using a Punnett square. Round, green and wrinkled, yellow offspring can also be calculated using the product rule, as each of these genotypes includes one dominant and one recessive phenotype. Therefore, the proportion of each is calculated as (3/4) × (1/4) = 3/16.
The law of independent assortment also indicates that a cross between yellow, wrinkled (YYrr) and green, round (yyRR) parents would yield the same F1 and F2 offspring as in the YYRR x yyrr cross.
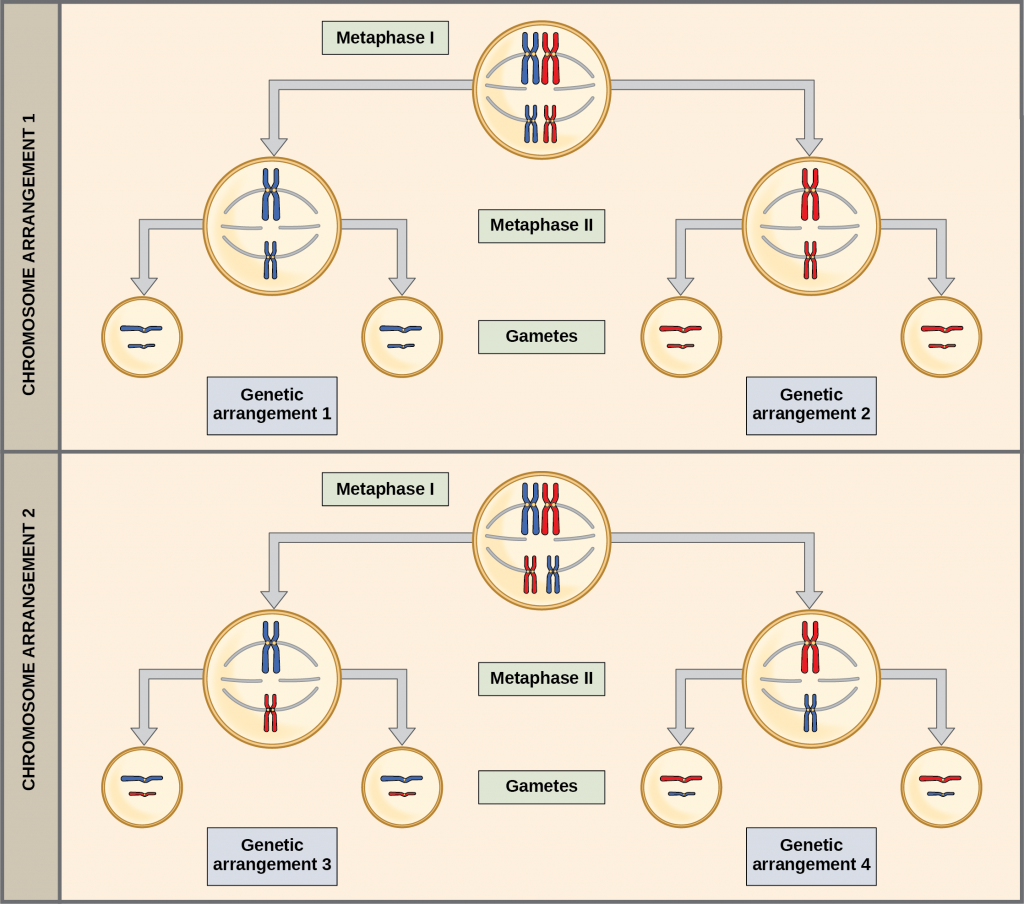
The physical basis for the law of independent assortment also lies in meiosis I, in which the two members of each homologous chromosome pair line up in random orientation. Each gamete will contain one copy of each chromosome type (sometimes the maternal copy and sometimes the paternal copy); hence, many different gamete types are possible based on different combinations of paternal and maternal chromosomes (and therefore the genes on them).
Identifying All Gamete Types Based on a Parental Genotype
To investigate the inheritance of traits, it is important to correctly predict all of the different gamete types that an individual of known genotype is expected to produce. Here, we need to remember that individuals are diploid and gametes are haploid. Hence individuals most often contain two copies of each gene (an exception is that human males only contain one copy of each X-linked gene) but gametes have only one copy of each gene. Because the segregation of alleles is random, half of the gametes are expected to contain one copy of the gene and the other half the other copy. When the assortment of genes is independent, either copy of gene 2 is equally likely to end up with either copy of gene 1.
What gamete types is an individual of genotype AabbCc expected to produce? Each gamete is expected to have one copy of gene A, one copy of gene B, and one copy of gene C. For gene A, each gamete would contain either the “A” or the “a” allele. Further, each gamete will contain a copy of “b” because that is the only allele for gene B in this individual. Since this individual is heterozygous for gene C, the same outcome is expected as for gene A: half of the gametes will contain the “C” allele and the other half will contain the “c” allele. An organized method to determine gamete types is to build a tree diagram in which each column represents a different gene.
Linked Genes
Although all of Mendel’s pea characteristics behaved according to the law of independent assortment, we now know that some allele combinations are not inherited independently of each other. Genes that are located on separate non-homologous chromosomes will always sort independently. However, genes on the same chromosome do not always sort independently. Linkage means that genes that are located physically close to each other on the same chromosome are more likely to be inherited together. However, because of the process of recombination, or “crossover,” it is possible for two genes on the same chromosome to behave independently, or as if they are not linked. To understand this, let’s consider the biological basis of gene linkage and recombination.
Homologous chromosomes possess the same genes in the same linear order. The alleles may differ on homologous chromosome pairs, but the genes to which they correspond do not. In preparation for the first division of meiosis, homologous chromosomes replicate and synapse. At this stage, segments of homologous chromosomes exchange linear segments of genetic material. This process is called recombination, or crossover, and it is a common genetic process. Because the genes are aligned during recombination, the gene order is not altered. Instead, the result of recombination is that maternal and paternal alleles are combined onto the same chromosome. Across a given chromosome, several recombination events may occur, causing extensive shuffling of alleles.
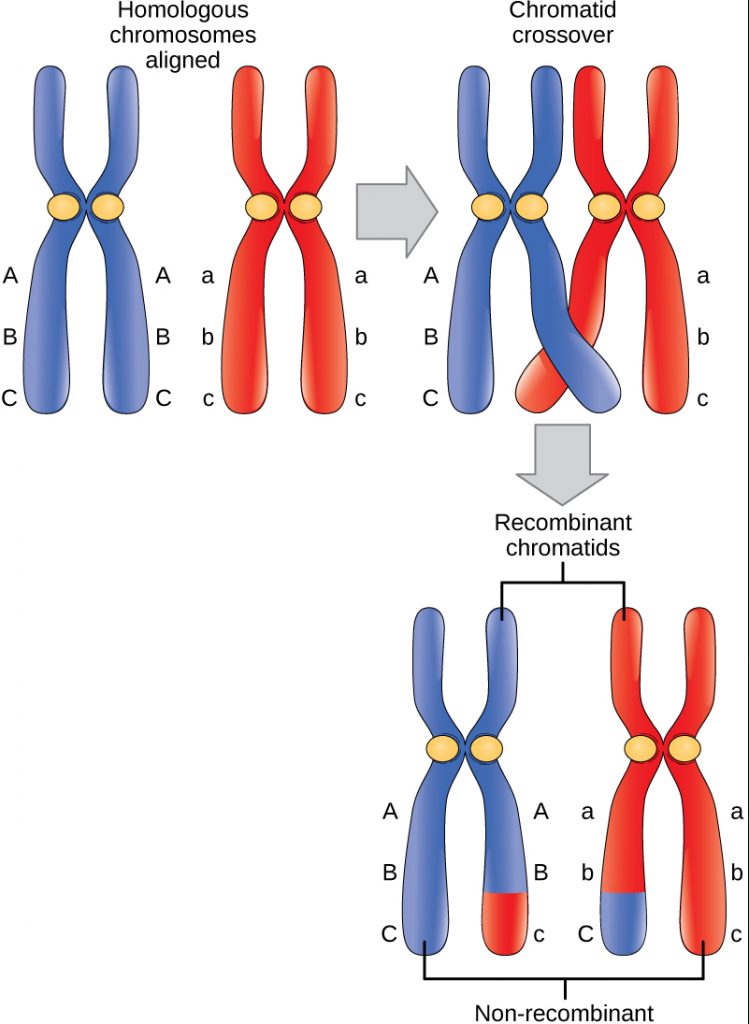
When two genes are located in close proximity on the same chromosome, they are considered linked, and their alleles, which are connected in the parental genome, tend to be transmitted through meiosis together.
On the diagram above representing a homologous chromosome pair, the “B” allele of gene B is linked to the “C” allele of gene C on one copy of the chromosome, and the “b” allele is linked to the “c” allele on the other copy. Gametes which retain this allelic arrangement are called parental or non-recombinant gametes (in this case BC and bc), and those in which the allelic arrangement is altered by crossover are called recombinant gametes (in this case Bc and bC). Because linked genes are more likely to be transmitted together, parental gametes are produced at a greater frequency than recombinant gametes. Thus, one can deduce which allele of gene C is linked to the “B” allele in a BbCc parent by identifying which two gamete types are produced at a higher frequency than the other two .
As the distance between two genes increases, the probability of one or more crossovers between them increases, and the genes behave more like they are on separate chromosomes. Geneticists have used the proportion of recombinant gametes (the ones unlike the parents) as a measure of how far apart genes are on a chromosome. Using this information, they have constructed elaborate maps of genes on chromosomes for well-studied organisms, including humans.
first round of meiotic cell division; referred to as reduction division because the daughter cells have half the number of chromosomes as the parent cell
phenomenon in which alleles that are located in close proximity to each other on the same chromosome are more likely to be inherited together
exchange of genetic material between homologous chromosomes resulting in chromosomes that incorporate genes from both parents of the organism
