Introduction to Supergroup Archaeplastida
The Archaeplastida is a monophyletic lineage of eukaryotic photoautotrophs. They have plastids acquired by primary endosymbiosis with a cyanobacterium. Members of this group have supplied plastids by secondary endosymbiosis to other photoautotrophic eukaryotic lineages (Euglenids, Alveolates, and Stramenopiles).
Archaeplastida includes:
- Glaucophyta – glaucophytes
- Rhodophyta – red algae
- Viridiplantae
- o Chlorophyta – green algae
- o Streptophyta
- Charophyta – charophytes (includes some groups previously classified as green algae)
- Embryophyta – plants
Glaucophyta (Glaucophytes)
This is a small (about a dozen species) and obscure group of unicellular freshwater algae. They are of interest to evolutionary biologists in that their chloroplasts possess an outer layer of peptidoglycan that is interpreted as being a remnant of the peptidoglycan cell wall of the cyanobacterial endosymbiont. We will not be examining glaucophytes.
Rhodophyta (Red Algae)
This is primarily a marine group, although it contains a small number of fresh and brackish water species. There are about 7,000 named species. The oldest member of this clade is the fossil Bangiomorpha isolated from rocks dated to 1.2 billion years ago. Current and fossil red algae are reef builders, predating the corals in early Paleozoic reefs. The modern “coralline” red algae form a calcium carbonate layer on their cell walls, and are still significant reef contributors (you may observe coralline red algae in our saltwater tank in the third floor foyer). Modern red algae are unicellular to multicellular, with most filamentous or multicellular. Many larger multicellular species are familiar as the reddish to blackish “seaweeds” (note: seaweed is a descriptive term that is not systematically useful. Other seaweeds include some of the green algae – Chlorophyta, while the most prominent seaweeds are usually kelps and other brown algae – Phaeophyta).
The most distinctive feature of red algae is their red-purple secondary photosynthetic pigments, produced by the reddish to bluish phycobilin pigments (phycocyanin, allophycocyanin, and phycoerythrin). These are the same secondary photosynthetic pigments found in cyanobacteria. Their reddish pigments allow red algae to live in deeper waters than most photoautotrophs. Red algae have a glycogen-like storage polysaccharide (floridean starch) and cellulosic cell walls. The cell walls of some red algae are the source of the commercially useful colloidal compounds agar or carrageenan. Red algae are used in some traditional cuisines including the familiar nori (Porphyra) that has been cultivated as a food source for centuries in Japan.
Red algae have a complex alternation of generations (a variation on a sporic life cycle) with up to three distinct generations: a haploid gametophyte (with male and female individuals), a diploid carposporophyte generation, and a diploid tetrasporophyte generation. These three generations may be isomorphic (appearing similar) or heteromorphic (appearing different). The gametophyte is the sexual generation, forming the haploid gametes. The diploid carposporophytes and tetrasporophytes reproduce by forming spores (diploid carpospores and haploid tetraspores). Red algae do not form flagellated cells at any stage in their life histories.
Chlorophyta (Green Algae)
This is a morphologically diverse and widespread clade of organisms often recognized at the rank of phylum. It includes about 4,500 species that may be free-living, mutualistic, commensal, or parasitic. Mutualistic unicellular green algae may be found as phycobionts in lichens or in associations with animals such as corals. The green algae includes unicellular, filamentous, colonial, and multicellular members. Some are coenocytic (not subdivided into cells). Green algae are common members of marine, brackish water, fresh water, and terrestrial ecosystems. They may be found in hot springs or hypersaline environments. One species of unicellular green alga is responsible for the reddish snow sometimes seen in Utah. Most form flagellated cells that are biflagellate (although the number of flagella ranges from 0 to many). Flagellar position is apical. All three life cycles (gametic, zygotic, and sporic) may be found in this clade.
Biochemically the green algae are very similar to terrestrial plants and charophytes, and these three groups are often placed in a single (unranked) clade, the Viridiplantae. All have similar secondary photosynthetic pigments (chlorophyll b, carotenoids, and xanthophylls) that give all three groups a similar green color. All have cellulosic cell walls and use starch as an energy storage molecule. Viridiplantae is generally recognized as being monophyletic.
Charophyta (charophytes, stoneworts)
Charophytes are a group of mostly freshwater unicellular, filamentous or multicellular algae that are closely related to terrestrial plants. Some (Chara and Nitella) have striking macroscopic organization such that they may easily be mistaken for members of the plant kingdom. They are sometimes grouped with the green algae, but have some cytological features (such as the formation of a phragmoplast during cytokinesis) that show their close affinity with plants. Charophytes and land plants are generally placed together in the (unranked) clade Streptophyta. Charophytes usually form biflagellate sperm (similar structurally to those of terrestrial plants) and have a zygotic life cycle.
Plantae (terrestrial plants, embryophytes)
This is the most familiar and diverse group in this supergroup with over 300,000 species. This group includes flowering plants, conifers, ferns, and mosses. This clade is customarily placed at the rank of kingdom (Plantae), but has also been placed at lower ranks (e.g. division Embryophyta). This group is primarily terrestrial (with some marine and freshwater members that are secondarily aquatic). All have a sporic life cycle, waxy spores (or pollen) that are waterproofed with sporopollenin, and an embryo that develops within a sterile jacket of cells (hence embryophytes).
Plants are one of the two most familiar groups of organisms (the other, of course, being the animals). More than any other clade, our concept of “what is a plant” has changed over the course of the last century. By the mid twentieth century the algal groups (“thallophytes”), fungi, and bacteria were placed elsewhere, with the remaining group, the embryophytes (originally the class Embryophyceae) being the sole recipients of the title “plant.” For the last five decades the embryophytes have been placed in their own kingdom (Plantae). The embryophytes nest within the Viridiplantae clade of the Archaeplastida. Their sister group within the Viridiplantae are the charophytes.
All plants have a sporic life cycle with multicellular sporophyte and gametophyte generations. The cycle is summarized below. Variations on this cycle are useful in the recognition and description of different plant clades.
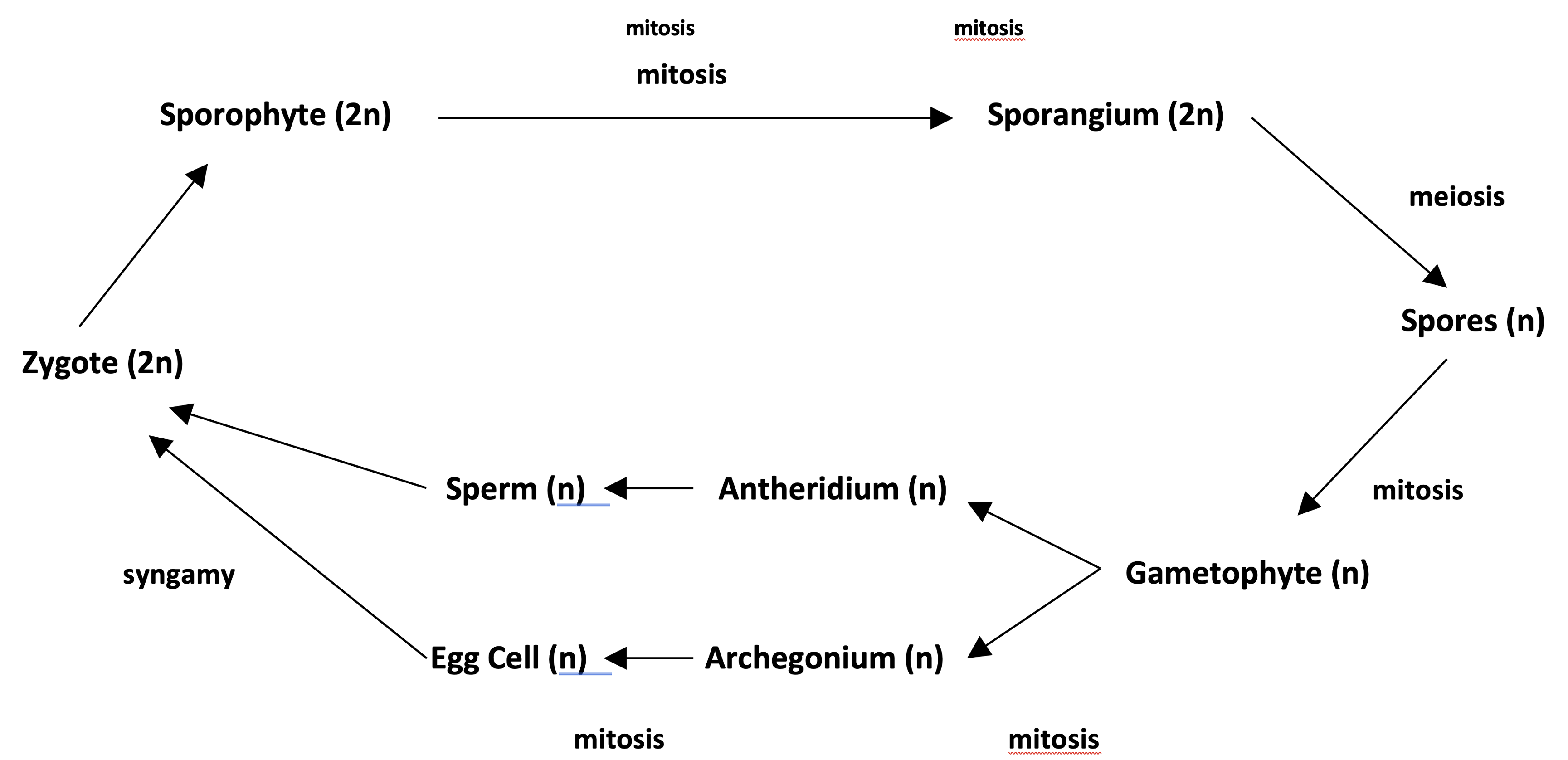
The plant kingdom is a terrestrial clade, and its most distinctive features are those that evolved during the transition to life on land. Key plant symplesiomorphies include:
- green pigments, cellulose cell walls, starchy polysaccharide energy storage molecules (these features shared with other Viridiplantae)
- sporic life cycle with specialized reproductive structures to protect reproductive cells from desiccation:
- o sperm and egg cells formed by the gametophyte within reproductive structures (gametangia) that have a “sterile jacket” of cells to protect the developing gametes.
- sperm formed within antheridia
- egg cells formed within archegonia
- o zygote develops into a young sporophyte (embryo) protected by the remnants of the multicellular archegonium of the gametophyte (hence embryophytes)
- o zygote and young sporophyte nourished by the gametophyte via placental transfer tissue
- o spores formed by the sporophyte within sporangia that have a sterile jacket of cells.
- spores (and pollen of seed plants) waterproofed with the complex waxy compound sporopollenin.
- formation of a waxy cuticle to reduce water loss through the epidermis
- formation of stomata for gas exchange through the cuticle (absent from liverworts)
- growth by an apical meristem or meristematic cell (a feature shared by some Viridiplantae)
- o sperm and egg cells formed by the gametophyte within reproductive structures (gametangia) that have a “sterile jacket” of cells to protect the developing gametes.
Derived features that occur within some (but not all) lineages of the plant kingdom include:
- vascular tissue (xylem and phloem) for rapid conduction of fluids (absent from mosses, liverworts, and hornworts)
- differentiation of the plant body into roots, stems, and leaves (absent from mosses, liverworts, and hornworts)
- reproduction by seeds (absent from the free-sporing plants: mosses, liverworts, hornworts, lycopods, and monilophytes)
The plant kingdom, as currently conceived, contains 10 clades recognized at the rank of phylum. These may be subject to a reduction in rank in future classifications:
- Kingdom Plantae (embryophytes)
- o Bryophyta (mosses)
- o Hepatophyta (liverworts)
- o Anthocerotophyta (hornworts)
- o Lycophyta (lycopods)
- o Monilophyta (ferns)
- o Pinophyta (conifers)
- o Cycadophyta (cycads)
- o Ginkgophyta (ginkgoes)
- o Gnetophyta (gnetophytes)
- o Magnoliophyta (flowering plants)
The first five phyla listed above have “free-sporing” life cycles in which the spores are released from the sporangia, and the gametophytes are (therefore) free-living. This lab is concerned with these divisions.
The second five divisions are seed plants in which the spores are retained (within sporophytes tissues) and develop into gametophytes within sporophyte tissue.
